
2026. 6. 22. · 00:22
Orientation selectivity: how V1 turns edges into signals
Orientation-selective neurons in V1 respond best to bars and edges at particular angles. This article explains the Hubel and Wiesel landmark experiments, the simple-cell edge-detector idea, modern debates about feedforward and cortical mechanisms, and why this concept matters for understanding how cortex builds visual features.

The problem: a line is not yet an object
Look at the edge of a table, a letter on a screen, or a branch crossing the sky. You experience it as part of a meaningful scene. In primary visual cortex, though, one of the first cortical questions is much simpler: is there a small light-dark boundary at this angle, in this tiny part of visual space?
That question is the heart of orientation selectivity. Many neurons in V1, the primary visual cortex, respond strongly when a bar, edge, or grating is tilted at one preferred angle, and much less when the same stimulus is rotated away from that angle. A vertical-edge cell is not a 「table」 cell. A 45-degree cell is not a 「tree branch」 cell. It is a local feature detector, one member of a much larger population that lets the visual system build shape from fragments.
Hubel and Wiesel's classic single-neuron recordings made this idea concrete. In 1959, they reported that neurons in the cat striate cortex had receptive fields with much richer structure than retinal or lateral geniculate nucleus cells: some cortical cells fired best for a line stimulus in a particular orientation and position, rather than for a simple spot of light. 1 Their 1962 paper then gave the field a vocabulary that still shapes visual neuroscience: simple cells, complex cells, end-stopped cells, receptive-field subregions, binocular interactions, and the functional architecture of orientation columns. 2
MIT 9.13 treats orientation selectivity as one of the gateway ideas in visual neuroscience because it shows how cortex changes the format of information. The retina and thalamus provide spatial contrast signals. V1 begins to represent local edges, contours, and texture elements, which are closer to the ingredients of seeing objects and scenes. 3
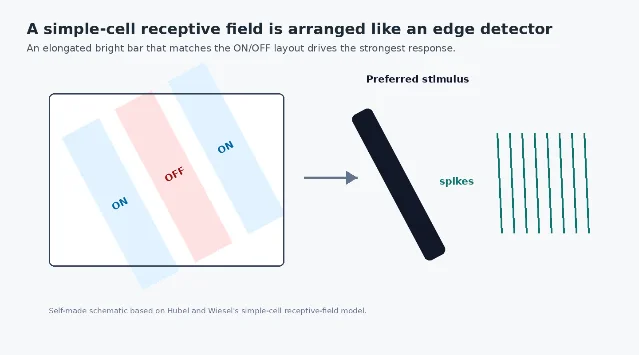
The landmark experiment: listening to one cortical neuron at a time
Hubel and Wiesel's experiments were simple in outline and radical in consequence. They recorded from single neurons in anesthetized cats while presenting visual stimuli on a screen. Instead of treating vision as a whole-field response, they asked what exact patch of visual space could drive one neuron.
A receptive field is that patch: the region of the visual field where a stimulus can change a neuron's firing. In retinal ganglion cells and many LGN neurons, receptive fields often look roughly circular, with center-surround organization. A light spot in the center might excite the cell, while light in the surround might suppress it, or vice versa. That design is excellent for detecting local contrast, but it is not yet a detector for a line at a particular slant.
In V1, Hubel and Wiesel found cells whose receptive fields were elongated and divided into subregions. For a simple cell, a narrow bar of light could excite the neuron if it lined up with the excitatory subregion, but rotating the bar by 90 degrees could make the same stimulus almost useless. The cell was not asking 「is there light here?」 It was asking 「is there contrast arranged in this orientation here?」 2
Their work also separated simple cells from complex cells. Simple cells have more clearly defined ON and OFF subregions: light in one part of the receptive field may excite, while light in a neighboring part may suppress. Complex cells are still orientation selective, but they are less fussy about the exact position of the stimulus within the receptive field. That difference hinted at a cortical hierarchy: outputs from simpler detectors could be combined to produce more tolerant feature responses.
This is why orientation selectivity became a model system for cortical computation. It is concrete enough to measure with a drifting bar or grating, but deep enough to raise general questions: how does cortex transform its inputs? How much selectivity is inherited from thalamus? How much is built by local cortical circuits? How does a population of such neurons tile the space of possible visual features?
Why one angle wins
The simplest cartoon begins with aligned inputs. Imagine several LGN neurons with circular center-surround receptive fields feeding into one V1 simple cell. If their receptive-field centers are arranged in a row, then a bar of light with the matching orientation activates many of them at once. Rotate the bar, and it no longer lines up with the row; fewer inputs arrive together, and the cortical neuron responds less.
That feedforward story captures the intuition of the Hubel-Wiesel model. It explains why a cortical neuron could become orientation selective even if its individual thalamic inputs are not strongly orientation selective. It also gives students a useful mental picture: V1 can detect a line by pooling several point-like contrast detectors in a particular spatial arrangement.
But the real circuit is not just a ruler made of LGN inputs. A modern review by Nicholas Priebe emphasizes that orientation selectivity marks a major representational jump from thalamus to cortex, while the mechanism remains a live question involving thalamic input, cortical excitation, inhibition, and intracellular response properties. 4 In other words, the feedforward arrangement is the starting hypothesis, not the whole answer.
One important experimental line asks how much of V1 orientation tuning depends on input from the lateral geniculate nucleus. Viswanathan and colleagues studied cat V1 and argued that feedforward geniculate inputs play an important role in generating orientation selectivity, while also showing that the cortical response cannot be reduced to a single static wiring diagram. 5 Other models show that recurrent cortical networks can sharpen, stabilize, or even generate selectivity under some conditions, especially through the balance of excitation and inhibition. 6
So the best answer to 「why does one angle win?」 is layered. The geometry of feedforward input matters. Local cortical circuitry matters. The neuron's membrane properties and spike threshold matter. The final tuning curve is the visible output of all those ingredients.
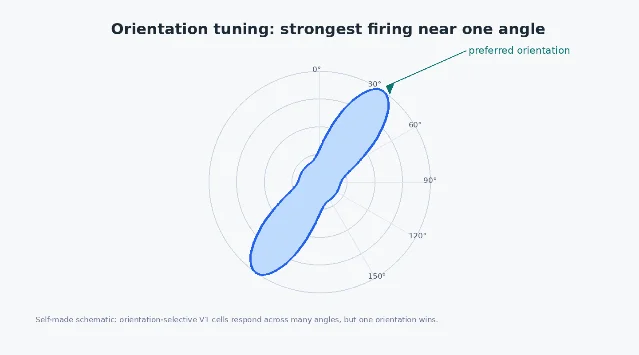
Tuning curves: not a yes-or-no detector
It is tempting to say that a V1 neuron 「detects vertical lines」. That is useful shorthand, but it hides an important point: orientation tuning is graded. A neuron might respond best to a vertical bar, somewhat to a bar tilted 15 degrees, weakly at 45 degrees, and hardly at all to a horizontal bar. The response profile across angles is its orientation tuning curve.
This matters because perception is population-based. A single neuron is noisy, limited, and local. The brain does not need one perfect vertical-line cell; it can read out a pattern across many neurons with different preferred orientations, positions, spatial scales, and response thresholds. Edges in the world are then represented by a distributed vote across a large set of tuned units.
The tuning-curve view also makes orientation selectivity experimentally useful. Researchers can quantify a neuron's preferred orientation, tuning width, response strength, contrast dependence, and invariance across position. Those measurements let scientists compare species, developmental stages, cortical layers, and computational models. A concept that began with bars of light on a screen became a precise way to ask how cortical circuits format sensory information.
From single cells to maps
Orientation selectivity does not exist only neuron by neuron. In many mammals, nearby V1 neurons with similar preferred orientations are arranged in orderly maps. In species such as cats and primates, orientation preference can rotate smoothly across the cortical surface, often around points called pinwheels, where many preferred orientations meet.
That map-like architecture was one of the reasons Hubel and Wiesel's work became so influential. Their 1962 paper did not only describe individual receptive fields; it helped launch the idea that cortex has a functional architecture, with columns and maps organizing what different populations of neurons compute. 2
Yet not all visual cortices use the same layout. Some rodents show a more 「salt-and-pepper」 arrangement, where neighboring neurons can prefer quite different orientations even though individual neurons are still orientation selective. Hansel and van Vreeswijk used this contrast to ask how orientation selectivity can arise without a smooth functional map. 6 More recent comparative work has found orientation pinwheels in the primary visual cortex of the tammar wallaby, a highly visual marsupial, showing that map organization can appear outside the standard cat-primate examples. 7
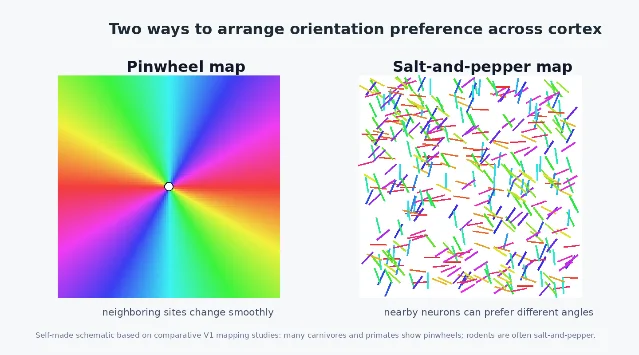
The map question changes how we think about V1. If orientation selectivity were only about single-cell tuning, one circuit solution might be enough. But if cortical tissue also arranges those preferences across space, we must explain two things at once: how a neuron becomes tuned, and why tuned neurons are placed where they are. Development, wiring constraints, visual experience, and evolutionary history all enter the story.
Why this small feature detector matters
Orientation selectivity is not the whole of vision. It does not explain face recognition, scene understanding, reading, attention, or visual consciousness. It is also not a license to say that the brain simply works like a modern convolutional neural network. The analogy between V1 filters and early convolutional filters is useful, but it should stay an analogy. Biological neurons spike, adapt, receive feedback, develop under genetic and sensory constraints, and sit inside recurrent circuits.
Still, orientation selectivity is one of the cleanest demonstrations that the cortex does not passively receive an image. It constructs a representation. Retinal signals are reorganized into features that are more useful for building contours, borders, surfaces, and eventually objects. That is why this concept belongs early in a cognitive neuroscience curriculum: it is a small window into a general principle.
The visual system solves big problems by decomposing them into many local questions. Is there contrast here? Is it arranged vertically, horizontally, or diagonally? Does this edge continue nearby? Do several fragments belong to the same contour? Later visual areas can use those answers to represent bodies, faces, places, words, and actions. But V1 orientation selectivity is one place where we can watch the transformation begin.
Next time you see a slanted line, try not to imagine that the eye hands the brain a finished picture. A better image is a crowd of V1 neurons, each voting most strongly for the tiny edge fragment that matches its preferred location and angle. Seeing starts, in part, as a chorus of local guesses.
Landmark paper
Hubel, D. H., & Wiesel, T. N. (1959). 「Receptive fields of single neurones in the cat's striate cortex.」 The Journal of Physiology. This paper established that V1 neurons can have structured receptive fields and selective responses that go beyond simple retinal center-surround coding. 1
Hubel, D. H., & Wiesel, T. N. (1962). 「Receptive fields, binocular interaction and functional architecture in the cat's visual cortex.」 The Journal of Physiology. This broader study introduced much of the vocabulary for simple and complex cells, orientation tuning, binocular interaction, and cortical functional architecture. 2
Course connection
This article connects directly to MIT 9.13 / Nancy's Brain Talks, 3.6 Orientation Selectivity, where orientation-selective V1 responses are used to introduce how primary visual cortex builds feature representations from earlier visual inputs. 3
참고 출처
- 1Hubel & Wiesel 1959, Journal of Physiology
- 2Hubel & Wiesel 1962, Journal of Physiology
- 3Nancy's Brain Talks, 3.6 Orientation Selectivity
- 4Priebe 2016, Annual Review of Vision Science
- 5Viswanathan et al. 2011, Journal of Neuroscience
- 6Hansel & van Vreeswijk 2012, Journal of Neuroscience
- 7Jung et al. 2022, Science Advances
이 콘텐츠를 둘러싼 관점이나 맥락을 계속 보강해 보세요.