
2026/6/23 · 0:28
Ocular dominance columns: why V1 keeps the eyes partly separate
Ocular dominance columns are alternating zones in primary visual cortex where one eye has more influence than the other. This article explains the Hubel and Wiesel discoveries, why early deprivation can reshape the map, and how newer adult-plasticity work complicates the classic critical-period story.

A strange thing happens early in the visual system. The two eyes do not simply send their signals into a common blender. In the primary visual cortex, many inputs are still tagged by eye of origin. Neighboring patches of cortex are biased toward the left eye or the right eye, forming alternating bands known as ocular dominance columns.
That may sound like a wiring detail, but it became one of the classic clues that cortex is organized as a map: not only a map of visual space, but also a map of which input channel has more influence. It also gave neuroscientists a concrete way to study a harder question: how early experience can reshape the brain.
What ocular dominance columns are
Ocular dominance columns are alternating zones in primary visual cortex, especially in layer 4C of striate cortex, where geniculocortical inputs serving one eye are more strongly represented than inputs serving the other. A cell can still be binocularly influenced; 「dominance」 does not mean the other eye is absent. It means one eye has more leverage over the response in that local patch of cortex. Adams and Horton summarize the anatomical idea this way: in some mammals, the afferents serving each eye segregate in layer 4C into stripes called ocular dominance columns. 1
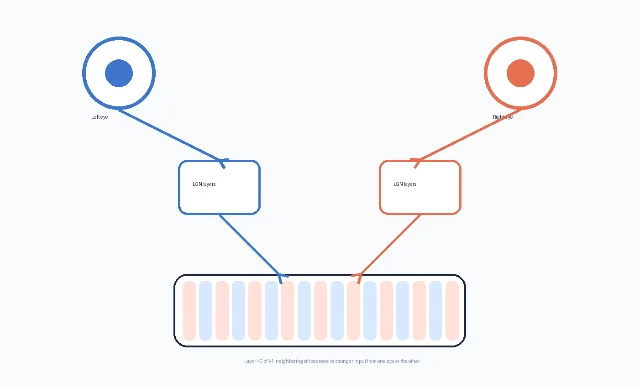
Two points help prevent confusion. First, ocular dominance columns are not the same as the visual field map. Retinotopy tells you where in visual space a patch of cortex is looking; ocular dominance tells you which eye has more weight there. Second, the columns are not simply 「left eye neurons」 versus 「right eye neurons」. They are part of a larger cortical architecture where eye preference, orientation preference, cytochrome oxidase patches, and local circuits overlap but do not reduce neatly to one another. 2
The landmark experiment: from single cells to cortical architecture
The classic foundation is Hubel and Wiesel's 1962 Journal of Physiology paper, 「Receptive fields, binocular interaction and functional architecture in the cat's visual cortex」. The paper is remembered for showing that neurons in cat primary visual cortex have structured receptive fields and that many cells respond best to oriented bars or edges. But the title's middle phrase, 「binocular interaction」, matters just as much for ocular dominance: the authors classified cells by how strongly each eye could drive them, creating the physiological language of eye dominance. 2
The move was powerful because it linked three scales. At the smallest scale, an electrode recorded the response of one neuron. At the middle scale, many neurons could be grouped by response properties such as receptive-field position, orientation, and eye preference. At the largest scale, cortex began to look like a repeating functional architecture rather than a homogeneous sheet. Ocular dominance columns became one of the clearest examples of that principle.
For a learner, the important conceptual jump is this: V1 is not just an image receiver. It is a structured workspace where different dimensions of the image and the input pathway are laid out in a partially ordered way. The columns are a map of competition and cooperation between the two eyes.
Why the columns mattered for critical periods
Ocular dominance columns became famous not only because they reveal cortical organization, but because they can be altered by early visual experience. Hubel and Wiesel's 1970 paper tested what happens when one eye is closed during development. They found a sharply time-limited window in kittens: susceptibility began near the start of the fourth week, stayed high between the sixth and eighth weeks, declined afterward, and disappeared around the end of the third month. Closing one eye in adult cats for more than a year produced no detectable physiological effect in their measurements. 3
That result is one reason the phrase 「critical period」 became central in neuroscience. A brief imbalance in input did not merely change a temporary percept. During the sensitive window, it shifted which eye could drive cortical cells. In Hubel and Wiesel's account, a few days of closure in the fourth week sharply reduced binocular influence and reduced the relative influence of the closed eye; a six-day closure could leave only a small fraction of striate cortex cells drivable by that eye. 3
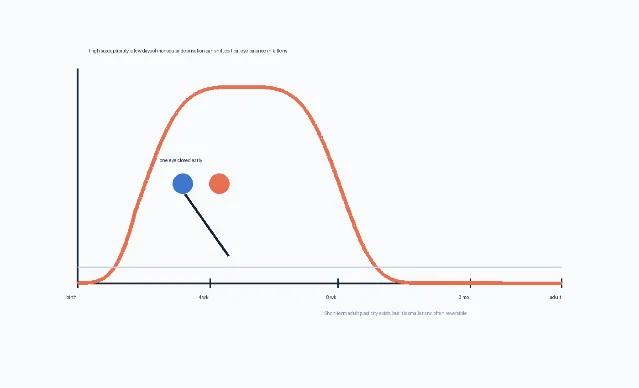
Later work in primates made the clinical stakes clearer. Horton and Hocking studied macaque striate cortex after monocular eyelid suture starting at different ages: 1, 3, 5, 7, and 12 weeks. Suture at 1 week caused the most severe shrinkage of deprived-eye columns; effects declined with later starting ages, and suture starting at 12 weeks caused no column shrinkage. The authors concluded that macaque V1 is most vulnerable to monocular deprivation during the first weeks after birth, supporting the clinical urgency of treating congenital cataract early. 4
This is the teaching power of ocular dominance columns. They turn an abstract developmental claim into a visible cortical pattern: early imbalance between the eyes can leave a structural and physiological footprint.
What the stripe map does not prove
It is tempting to say that ocular dominance columns exist so that the brain can compute depth from the two eyes. The evidence does not let us say that so cleanly. Adams and Horton emphasize that the function of ocular dominance columns remains unresolved. Some species with stereopsis lack ocular dominance columns, so the columns cannot be strictly required for stereoscopic vision. They also are not required for the existence of monocular cells in layer 4C. 1
The cross-species pattern is surprisingly uneven. Adams and Horton list ocular dominance columns in cats, ferrets, macaques, humans, and several other primates, but not in rats, mice, rabbits, sheep, goats, and some other mammals. Squirrel monkeys are especially revealing: some individuals have clear columns, some have essentially none, and some show columns only in part of striate cortex. 1

Even within species, the map is not a fixed barcode. In humans and macaques, column patterns can vary substantially across individuals; in normal macaques, Adams and Horton report at least a twofold range in column width. This variability is not a nuisance detail. It is part of the scientific puzzle. A cortical feature can be robust enough to be named and studied, yet variable enough that its function and developmental rules remain open.
Are the columns built by experience?
The deprivation experiments might make it sound as if visual experience builds ocular dominance columns from scratch. That is too simple. The review evidence points in the opposite direction for initial formation: in macaques, an adult-like pattern is already well formed at birth, before visual experience, and column formation begins late in gestation. In ferrets, initial formation can occur even after binocular enucleation at birth, meaning visual experience is not required for the first segregation. 1
A better formulation is: early biological programs and spontaneous activity can set up the initial map, while visual experience can refine, rebalance, or distort it during sensitive windows. This distinction matters because it avoids a false nature-versus-nurture story. The columns are neither purely hardwired nor freely sculpted by experience at any age. They sit at the intersection of development, competition, and use.
This is also why ocular dominance became such a useful model system. If you want to study how genes, spontaneous neural activity, sensory input, and synaptic competition interact, the two eyes provide a natural experiment. They deliver related but separable streams into the same cortical area.
What adult plasticity changes in the story
The classic critical-period story should not be read as 「the adult visual cortex cannot change」. It means that the large developmental remodeling seen after early deprivation is strongly age-dependent. Recent work on short-term monocular deprivation in adults has reopened the question of how flexible eye balance remains after development. Min's 2025 mini-review summarizes a line of psychophysical, physiological, and neuroimaging studies showing that several hours of depriving one eye can temporarily shift perceptual dominance toward the deprived eye in adults. 5
The cautious word is temporarily. Min notes that a single short-term deprivation session in adults produces a transient effect, often lasting on the order of tens of minutes, and that the evidence for boosting or prolonging such plasticity is mixed. The same review discusses possible relevance to amblyopia treatment, including inverse patching approaches, but it also stresses limits: short-term monocular deprivation alone does not fully restore binocular balance, and responses vary across people and clinical conditions. 5
So the modern story is more precise than the old slogan. Early development is special, but adult cortex is not inert. The question has shifted from 「plastic or not」 to 「what kind of plasticity, on what timescale, through which mechanisms, and with what clinical effect?」
Why this concept matters
Ocular dominance columns matter because they make cortical organization concrete. They show that cortex can be divided into repeating local territories, each biased toward a particular input channel, while still participating in a shared perceptual task. They also show why development is not merely growth in size. During a sensitive period, the balance of input can alter which pathways get cortical territory.
For cognitive neuroscience, this concept is a bridge. It connects the single-neuron world of receptive fields to the systems-level world of brain maps. It connects developmental neuroscience to clinical problems such as cataract and amblyopia. And it gives the MIT 9.11 / Nancy's Brain Talks sequence a useful early visual-cortex example before moving into higher-level visual regions: functional organization begins long before faces, places, and words. The course page for 「3.7 – Ocular Dominance Columns」 sits inside a broader course that emphasizes functional organization, development, specificity, and the limits of inference from methods. 6
If orientation selectivity taught us that V1 extracts local edges, ocular dominance columns teach a complementary lesson: V1 also preserves and organizes the history of input. The cortex does not erase the two eyes as soon as signals arrive. It keeps their rivalry and cooperation visible in its own architecture.
Landmark paper: Hubel, D. H., & Wiesel, T. N. (1962). 「Receptive fields, binocular interaction and functional architecture in the cat's visual cortex」. The Journal of Physiology. DOI: 10.1113/jphysiol.1962.sp006837. 2
Course connection: Nancy Kanwisher's MIT Brain Talks page 「3.7 – Ocular Dominance Columns」 places this concept in the early visual-cortex part of the course, before the sequence moves into higher-level visual recognition and other cognitive systems. 6
参考ソース
- 1Ocular Dominance Columns: Enigmas and Challenges
- 2Receptive fields, binocular interaction and functional architecture in the cat's visual cortex
- 3The period of susceptibility to the physiological effects of unilateral eye closure in kittens
- 4Timing of the Critical Period for Plasticity of Ocular Dominance Columns in Macaque Striate Cortex
- 5Ocular Dominance Plasticity: A Mini-Review
- 63.7 – Ocular Dominance Columns

このコンテンツについて、さらに観点や背景を補足しましょう。