Border Cells: How the Brain Marks the Edges of Space
Border cells fire when an animal is near an environmental boundary, giving the brain's navigation system an anchor for walls, edges, barriers, and drops. This article explains the 2008 Solstad et al. landmark paper, the boundary vector cell model, and why boundaries help stabilize place and grid maps.

Border cells answer a simple navigation problem that pure coordinates do not solve: where are the edges? A grid cell can tile an open floor. A head direction cell can say which way the head points. A place cell can mark one patch of space. But an animal also needs to know where the wall, drop, barrier, or doorway is, because boundaries make a map usable.
The best short definition is this: a border cell fires when the animal is close to a boundary at a particular side of the local environment. In the 2008 landmark paper, Solstad, Boccara, Kropff, Moser, and Moser described neurons in the medial entorhinal cortex whose activity hugged environmental borders and stayed tied to those borders when boxes were stretched or changed in size and shape. 1
That makes border cells the fourth member of the navigation set we have been building: place cells in the hippocampus, grid cells and head direction cells in medial entorhinal cortex, and now border cells, which tell the system how the map is anchored to the geometry of the surrounding world. 2
What a border cell is
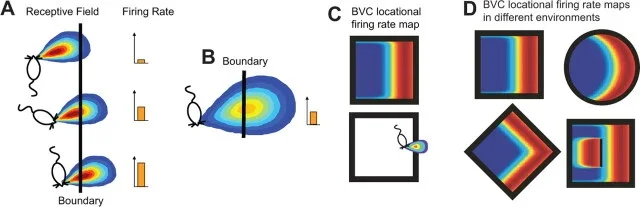
A border cell does not fire everywhere near every wall. It has a preference. One cell may fire along the north wall of a square box. Another may fire along the east wall. Another may respond near a corner because two borders satisfy its preferred geometry. The firing is spatial, but it is spatial in a different way from place cells.
Place cells usually mark a small location: roughly, "here." Grid cells mark many locations in a repeating triangular lattice: roughly, "this coordinate phase." Border cells mark a relation between the animal and the edge of an environment: roughly, "near this kind of boundary in this direction." Solstad and colleagues reported that these cells were relatively sparse, less than 10% of the local population, and appeared in medial entorhinal cortex and nearby parasubiculum, intermingled with grid cells and head direction cells. 1
The anatomical location matters. The medial entorhinal cortex is a major input and output hub for the hippocampus. If place cells are the hippocampus's location code, the entorhinal cortex supplies several kinds of structure that can help build that code: metric spacing from grid cells, orientation from head direction cells, and environmental shape from border cells. 2
The landmark experiment
The 2008 Science paper searched for neurons that fired when rats explored environments with different walls and shapes. The critical test was not just whether a neuron fired near a wall in one box. Many spatial neurons can appear wall-related by accident. The stronger test was whether the activity stayed attached to the appropriate geometric border when the environment was stretched or when the animal moved through enclosures of different size and shape.
The result was the signature pattern: orientation-specific, edge-apposing activity. A cell could remain active near the same type of border after changes to the enclosure. The abstract of the paper is unusually direct: the authors reported a cell type that fires when an animal is close to the borders of the proximal environment, and they proposed that these cells could help anchor grid fields and place fields to a geometric reference frame. 1
That last phrase is the point. Border cells are not a decorative add-on to the grid system. They solve the problem of drift. A self-motion-based coordinate system can accumulate error; a boundary gives the map something in the world to lock onto.
Why theorists expected them first
Border cells were not discovered in a vacuum. A family of models had already argued that place cells might be built partly from inputs that encode the vector from the animal to environmental boundaries. Barry, Lever, Hayman, Hartley, Burton, O'Keefe, Jeffery, and Burgess reviewed this boundary vector cell model in 2006. In that model, a boundary vector cell fires best when a barrier lies at a particular distance and allocentric direction from the animal. 4
The model was attractive because it explained a classic puzzle. When an experimental box is stretched, some place fields stretch with it, split, or stay attached to nearby walls. A pure coordinate code has trouble with that. A boundary-based input makes it natural: change the walls, and the inputs that depend on those walls change too.

A few years later, Lever and colleagues recorded boundary vector cells in the subiculum, a hippocampal formation structure downstream of CA1. They reported cells whose firing fields matched the predicted description: the cell responded to boundaries at a preferred distance and allocentric direction, and the pattern was much more about the animal's location relative to a boundary than about which way the rat was facing. 3
This gives the field two closely related ideas. Border cells usually refers to the medial entorhinal and parasubicular cells described by the Moser group. Boundary vector cells refers to the broader computational and physiological idea that neurons can encode boundaries at a specific distance and direction. The two are not always treated as identical, but they point to the same principle: geometry is coded directly, not inferred only after place cells fire.
What boundaries add to a cognitive map
A useful map needs at least two ingredients. It needs a coordinate-like code that can support movement through open space, and it needs anchors that keep that code aligned to the external world. Grid cells are excellent candidates for the coordinate-like part. Border cells are excellent candidates for the anchor part.

This is why border cells matter for place cells. A hippocampal place field often sits in a location defined by distances to walls: a corner, a region near one side, or a point whose position changes when the room's shape changes. Boundary vector models can produce place fields by combining several boundary-tuned inputs. In plain language, the place cell can say "fire at this location" because its inputs say "the north wall is here, the east wall is there, and the animal is at the intersection implied by those distances." 4
The same logic helps explain why boundaries dominate navigation. A chair can move. A smell can fade. A visual cue card can be rotated by an experimenter. But walls and edges define the reachable shape of the world. In the Lever 2009 study, boundary-responsive firing could follow walls, circular enclosures, platform edges, and even drops, showing that the relevant signal is not merely "visual wall" but a more abstract boundary of accessible space. 3

What is still debated
The main debate is not whether boundary-related neurons exist. They do. The hard questions are about function and hierarchy.
One question is whether border cells generate place fields, stabilize them, or both. Boundary vector models show that boundary inputs can help explain place-cell geometry, but real hippocampal maps also depend on grid cells, sensory cues, head direction signals, path integration, reward, task demands, and memory history. A border signal is a powerful input, not the whole map.
Another question is how to relate medial entorhinal border cells to subicular boundary vector cells. The 2009 subiculum paper explicitly discusses the relationship between those cells and medial entorhinal border cells, rather than treating them as the same population. 3 The careful version is: the brain has multiple boundary-sensitive signals, and different parts of the hippocampal-entorhinal system may use them for different operations.
A third question is how abstract the boundary code is. Does it respond to walls, cliffs, doorways, route limits, invisible task boundaries, or event boundaries in memory? The strongest evidence here comes from physical space. Extending the idea into abstract cognition is tempting, but it should stay marked as an extrapolation unless the experiment directly tests it.
Why this concept matters
Border cells make the cognitive map less like graph paper and more like a map of a real room. The brain does not only compute where you are in a coordinate system. It also represents the edges that make the coordinate system meaningful.
That is why this concept belongs after place cells, grid cells, head direction cells, and retrosplenial cortex. Place cells say where. Grid cells supply a metric. Head direction cells say which way. Retrosplenial cortex helps connect egocentric views with allocentric maps. Border cells add the environmental frame: the walls, edges, and barriers that make a place a place.
Landmark paper: Solstad, Boccara, Kropff, Moser, and Moser, "Representation of geometric borders in the entorhinal cortex," Science, 2008. 1
Course connection: MIT 9.13's navigation sequence asks how the human brain represents places, scenes, and routes. Border cells fit naturally beside Lecture 9, which continues the course's treatment of scene perception and navigation. 5
围绕这条内容继续补充观点或上下文。