Cooking oil at 10% of your calories suppresses EPA. A new RCT shows exactly how.
Sergeant et al. (2026, Nutrients, published June 4) conducted a 12-week double-blind controlled-feeding RCT in 52 healthy adults and found that a high-LA diet (10% of energy, typical of most Western seed-oil use) selectively suppressed plasma EPA and oxylipin balance — providing the first direct human controlled-feeding evidence for LA–EPA substrate competition at the shared FADS1 enzyme.

研究速览
Seed oils and "omega-6 overload" generate enough online heat to qualify as their own food fight. Most of that argument has moved faster than the human clinical data behind it. A double-blind controlled-feeding trial published June 4, 2026 in Nutrients now adds something the debate has largely lacked: direct, prospectively measured evidence in humans showing that raising dietary linoleic acid (LA) from 2.5% to 10% of daily calories selectively reduces plasma EPA — the anti-inflammatory n-3 fatty acid — without meaningfully changing the n-6 end-product, arachidonic acid (ARA).
The study's headline result is mechanistic, not clinical. The authors do not report cardiovascular events, inflammation scores, or disease outcomes. What they show is how much the biosynthetic balance shifts at the enzyme level when LA intake reflects a typical Western diet — and what that shift does downstream, in the cells that manage inflammatory signaling. 1
Study design and population
Study type: Double-blind, randomized, parallel-arm controlled-feeding trial · Registry: NCT02962128 · Journal: Nutrients (MDPI, open access CC BY 4.0) · Published: June 4, 2026 · Peer-review status: Published (submitted April 1; accepted May 27, 2026)
Lead institution: Wake Forest University Health Sciences Clinical Research Unit, Winston-Salem, NC. Senior and corresponding author Floyd H. Chilton is a co-founder of Resonance Pharma, Inc. (a company developing phospholipase diagnostics and therapeutics), a relationship managed through the University of Arizona. No other authors declared conflicts of interest. Funding came from NIH grant R01 AT008621 (National Center for Complementary and Integrative Health) and the Wake Forest Clinical and Translational Science Award (NIH UL1TR001420). 1 2
Participants: 80 healthy adults (age 21–65, mean ~38; 63–71% female; predominantly European ancestry) with BMI 19–30 and no major metabolic disease. People taking NSAIDs, steroids, or fish-oil supplements were excluded. Baseline plasma fatty acid profiles were comparable between arms; only HDL-C (56.3 vs. 65.5 mg/dL, p = 0.007) and fasting glucose (92.5 vs. 87.6 mg/dL, p = 0.002) differed at randomization.
Intervention: Participants were randomized to 12 weeks of controlled feeding, with over 95% of daily fat calories prepared and provided by a metabolic kitchen. The Low-LA arm ate a diet containing 2.5% of energy from LA, achieved with high-oleic safflower oil (~13% LA). The High-LA arm ate a diet with 10.0% of energy from LA, achieved with high-linoleic safflower oil (~73% LA). Alpha-linolenic acid (ALA, the plant-based n-3 precursor) was held constant at ~1% of energy from flaxseed oil in both arms. Total fat was 25–30% of calories in both conditions, and total caloric intake was matched. Participants met with a registered dietitian weekly for food pickup and counseling.
Sixteen of 80 participants withdrew (20%), primarily from the Low-LA arm, mostly due to scheduling and adherence constraints. The final analysis included 52 completers (25 Low-LA, 27 High-LA). 1
Primary outcomes: Plasma concentrations of LA, ARA, EPA, and DHA at 12 weeks. Secondary outcomes included 16 additional fatty acid species and a profile of 40 oxylipin species generated by zymosan-stimulated whole-blood ex vivo assay, measured via liquid chromatography–tandem mass spectrometry (LC-MS/MS) at weeks 0, 4, 8, and 12.
What high LA does to EPA — and why ARA is protected
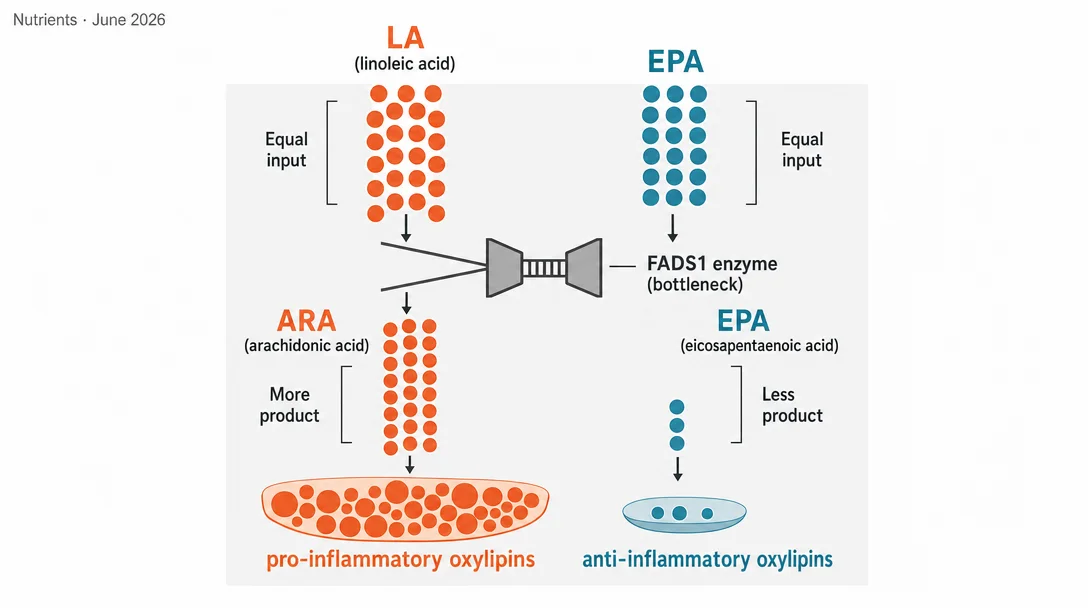
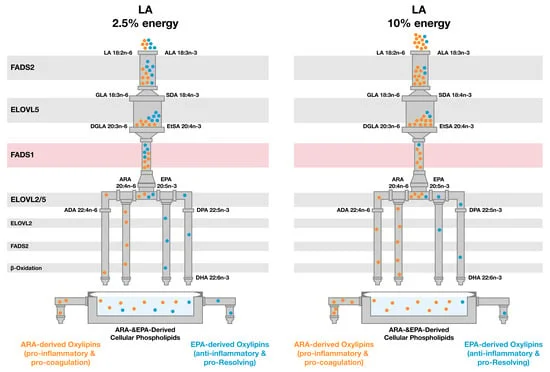
The trial's central finding turns on a structural feature of fatty acid metabolism: the n-6 and n-3 pathways share the same elongase and desaturase enzymes, starting with FADS2 (Δ6-desaturase) and converging at FADS1 (Δ5-desaturase). ARA is the n-6 end-product; EPA is the n-3 end-product. When dietary LA is abundant, it floods the early steps of the pathway, forcing n-3 precursors to compete for the same enzyme capacity.
At 12 weeks, plasma EPA and ETA (another n-3 highly unsaturated fatty acid, or HUFA) were both markedly lower in the High-LA arm compared to Low-LA. The statistical relationship between plasma EPA and ARA across the intervention period was strongly LA-dependent (F = 27.85, p < 0.0001), while the DHA-ARA relationship was far weaker (F = 4.48, p = 0.036). 1
What makes this mechanistically informative is the ARA result: plasma ARA did not differ between the two arms. The authors interpret this as saturation of the n-6 desaturation pathway — approximately 2.5% of energy from LA is already enough to fill ARA production close to its ceiling. Pushing LA intake to 10% (a level typical of Western diets dominated by soybean, corn, and sunflower oils) does not increase ARA further; it simply competes more aggressively for the shared enzymes, pulling substrate away from EPA synthesis.
DGLA (dihomo-gamma-linolenic acid), an n-6 intermediate that sits just before the FADS1 gate, was slightly but significantly elevated in the High-LA arm — consistent with increased flux through the early elongation steps without a matching increase at the Δ5-desaturase step. This pattern points to FADS1 capacity as the rate-limiting constraint under high LA conditions.
DHA showed only a non-significant trend toward reduction. The authors suggest that multiple additional biosynthetic steps and a preferential conservation mechanism may buffer DHA against LA competition more effectively than EPA. As they write, "the selective suppression of EPA, while DHA showed a non-significant trend toward reduction, identifies EPA as the most diet-responsive node in the endogenous n-3 pathway under competitive LA exposure." 1
The ARA/EPA ratio was significantly elevated in the High-LA arm at every measured time point. The total n-6/n-3 HUFA ratio was also markedly higher, driven by both the elevated n-6 substrate load and the suppressed n-3 HUFA production.
The downstream consequence: oxylipin balance
Plasma fatty acid ratios are a biomarker. What happens at the signaling level is measured by the oxylipin assay — the study's most functionally relevant outcome.
Oxylipins are short-lived lipid mediators produced when immune cells process fatty acids in response to a stimulus. They regulate inflammation onset, intensity, and resolution. ARA-derived oxylipins include prostaglandins and leukotrienes associated with pro-inflammatory signaling; EPA-derived oxylipins tend toward anti-inflammatory and pro-resolving functions. The two families are made by the same enzymes — 5-lipoxygenase (5-LOX) processes whichever substrate is most available.
The Sergeant et al. team used a zymosan-stimulated whole-blood assay: they took blood from participants, challenged it with zymosan (a fungal cell-wall particle that triggers leukocyte 5-LOX activity), and quantified 40 oxylipin species via LC-MS/MS. This captures integrated leukocyte enzymatic capacity under a standardized stimulus — not static circulating oxylipin levels, but the functional output of the inflammatory machinery.
The 5-HETE/5-HEPE ratio (ARA-derived / EPA-derived) was consistently higher in the High-LA arm across the intervention, as was the 5-HETE/5-HETrE ratio (ARA-derived / DGLA-derived). 1 Both ratios index substrate competition at 5-LOX directly: the enzyme uses ARA, EPA, and DGLA as substrates, and its output tilts toward whichever is more abundant. When EPA is suppressed by high dietary LA, 5-LOX processes proportionally more ARA.
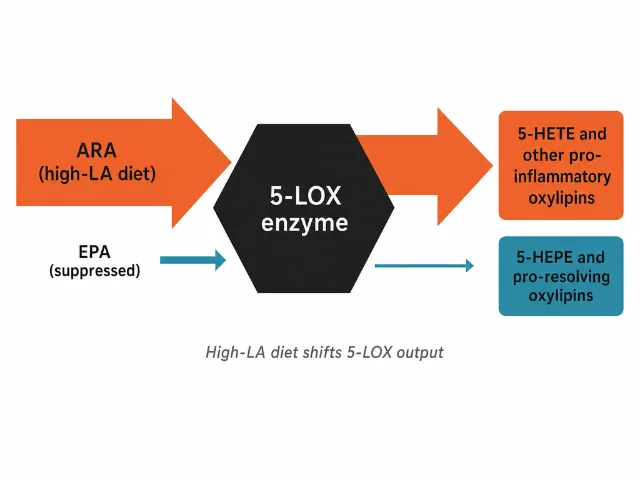
The authors are careful about interpretation: "these alterations have the capacity to systematically recalibrate the balance of pro- and anti-inflammatory eicosanoid signaling through substrate-level competition without requiring changes in total fat intake." The key qualifier is "capacity." The ex vivo assay confirms the enzyme is getting proportionally more ARA substrate under high-LA conditions — it does not confirm that this shift produces measurable changes in in vivo inflammation, let alone clinical outcomes.
正在加载内容卡片…
Limitations and what this study cannot tell you
Several constraints shape how far these results extend.
No clinical endpoints. This is the most important caveat, and the authors state it explicitly: "Whether sustained LA-driven flux redistribution influences long-term disease trajectories remains to be determined." Plasma EPA suppression and a shifted oxylipin ratio are biomarkers, not proxies for cardiovascular events, inflammatory disease incidence, or mortality. Studies that moved rapidly from this type of mechanistic observation to clinical claims — notably some of the Sydney Diet Heart Study re-analyses that the authors cite — have a mixed track record in replication. 1
Moderate sample size and 20% attrition. Eighty participants randomized, 52 completed. The attrition was heavier in the Low-LA arm, where replacing most dietary fat with high-oleic safflower oil proved harder to sustain. Exact plasma concentration values (mg/dL) for each fatty acid were not reported in the main text — they are in Supplementary Table S4, which was not accessible for independent verification.
The control condition uses oleic acid as the LA substitute. Both arms received different fatty acid profiles, not just different LA levels. The authors justify this by noting that oleic acid (a monounsaturated fat) is not a substrate for Δ6-desaturase and does not compete in the PUFA elongation-desaturation pathway — making it an appropriate non-competing substitute. That is scientifically defensible, but the two arms are not pure LA-dose comparison: they represent distinct dietary fat environments.
FADS1/FADS2 genotype was not accounted for. These gene variants substantially alter the efficiency of fatty acid desaturation and are distributed differently across ancestry groups. The trial was not powered for genotype stratification. The authors flag this as "a priority for future investigations" — individuals carrying FADS1 loss-of-function variants may respond very differently than the population mean. 1
Twelve weeks is short for long-term inference. The trial cannot speak to whether the EPA suppression effect persists, worsens, or adapts over years of a Western diet.
The oxylipin assay is ex vivo, not in vivo. Zymosan-stimulated whole-blood assays capture enzymatic capacity under controlled conditions; they do not directly reflect what happens inside tissues during an actual infection, injury, or chronic inflammatory process.
Dietary takeaway
This trial does not make a case for eliminating cooking oils or categorically avoiding polyunsaturated fats. What it establishes — for the first time in a controlled human trial — is that eating LA at the level common in Western diets produces a measurable, selective suppression of EPA biosynthesis, and that this suppression shifts functional oxylipin output toward ARA-derived mediators when immune cells are challenged.
The actionable reading depends on your starting point:
If you rely primarily on soybean, corn, or sunflower oil as your cooking fat, this study provides the first controlled human evidence that doing so likely suppresses EPA synthesis. You are not necessarily producing more ARA — the n-6 pathway appears saturated at low LA intakes — but you are crowding out the n-3 side. The practical options are: replace some of that oil with a lower-LA alternative (extra-virgin olive oil is predominantly oleic acid, the same fat used in the Low-LA arm here; avocado oil is similar), increase dietary EPA intake from fish or marine sources to compensate for reduced endogenous synthesis, or both.
If you already eat fatty fish two or more times per week or take a marine omega-3 supplement, the relevance of this study is lower. Dietary EPA bypasses the biosynthetic pathway entirely — it enters circulation directly without competing at FADS1. The substrate competition this trial documents operates specifically on the endogenous synthesis route from ALA.
For dietitians advising clients on fat quality: the framing this study supports is "LA and ALA compete for the same enzymes, and Western LA intakes push that competition hard against n-3 synthesis." That is now a statement grounded in a 12-week double-blind controlled-feeding trial rather than animal and cell data alone. "Reducing omega-6 intake" and "increasing omega-3 intake" are not fully interchangeable strategies — reducing LA addresses the substrate competition, while increasing EPA/DHA directly supplements the end-product. Both levers work at different steps.
The authors also raise a health disparities angle worth noting for practice: low-income populations in the US consume more ultra-processed foods formulated with high-LA seed oils and less fish and n-3-rich food overall, meaning the competitive EPA suppression documented here is most likely to be consequential in precisely those populations with the least dietary buffer against it. 1
One caution to hold throughout: these are biomarker results from 52 people over 12 weeks. Whether the EPA suppression and oxylipin shift seen here translate into measurable differences in inflammatory disease risk over years of a Western diet is the question this study raises without answering. The authors are explicit that they do not. That is not a limitation of the research — it is an honest representation of what mechanism studies can and cannot show.
Cover image: AI-generated editorial diagram.
围绕这条内容继续补充观点或上下文。