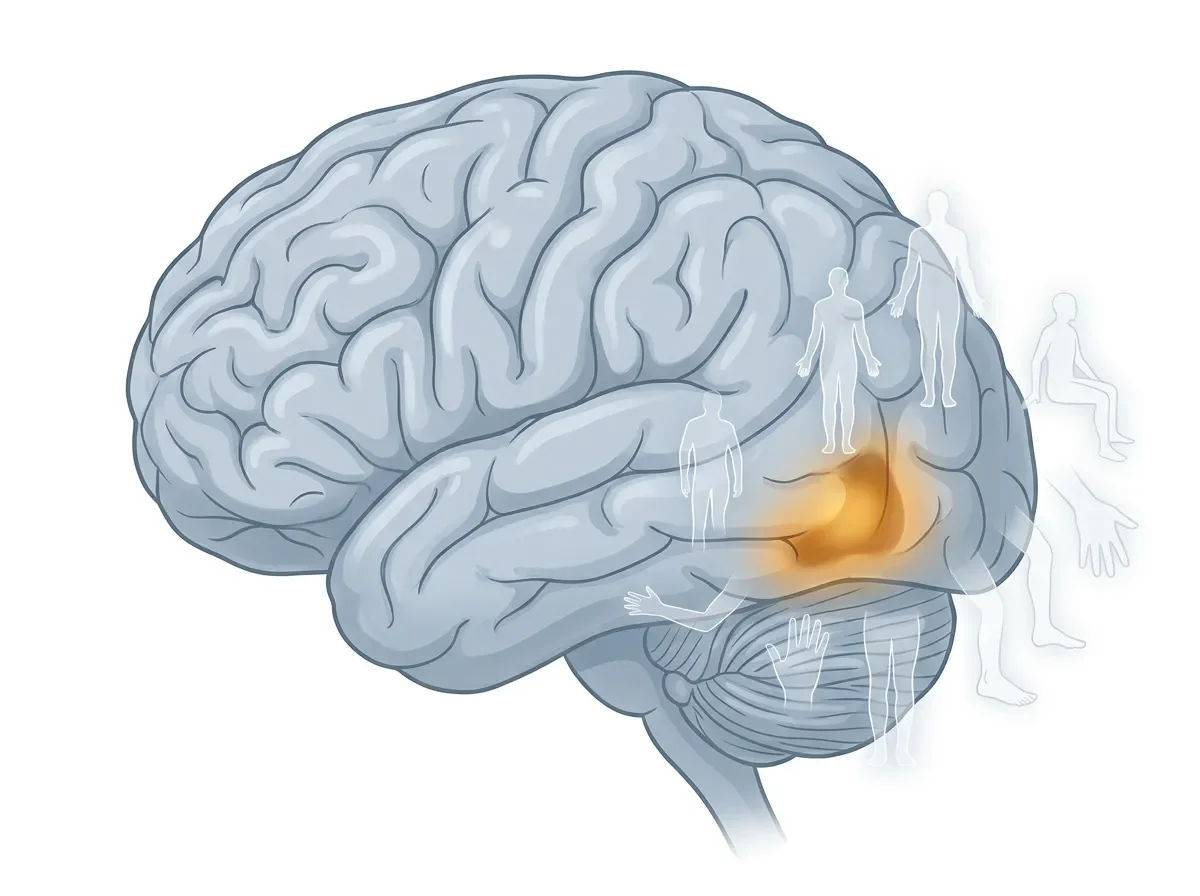
The Extrastriate Body Area: How the Brain Represents the Human Form
In 2001, Downing, Jiang, Shuman & Kanwisher discovered a patch of lateral occipitotemporal cortex that fires selectively to images of human bodies and body parts — the extrastriate body area (EBA). This article covers its location, what it responds to, what TMS disruption reveals about its causal role, how it relates to the nearby fusiform body area (FBA), and why its existence matters for theories of category-selective visual cortex.

The human visual system handles faces by dedicating a patch of cortex to them — the fusiform face area (FFA), which we covered in the first article in this series. The parahippocampal place area (PPA) does something similar for scenes and spatial layouts. The natural question, once both had been identified, was whether the same logic applied to the rest of the human form. Are bodies represented somewhere in particular, or is that information just distributed across general object-recognition circuits?
The answer, it turned out, was that bodies have their own address: a region in the lateral occipitotemporal cortex now called the extrastriate body area, or EBA.
The 2001 discovery
In a series of six fMRI experiments, Paul Downing, Yuhong Jiang, Miles Shuman, and Nancy Kanwisher showed that a bilateral region of lateral occipitotemporal cortex responded substantially more to images of human bodies and body parts than to a wide range of control stimuli — faces, objects, houses, scrambled images, and non-human animals 1. The pattern held up across participants, experiments, and control conditions. The region was present in every individual tested, somewhat more strongly expressed in the right hemisphere.
The stimuli in the key localizer contrasts were deliberately simple: grayscale photographs of headless human bodies (torsos, arms, legs) versus chairs or other object categories. By excluding faces from the body stimuli, the researchers isolated a body-selective response that was genuinely distinct from the face-selective response of the FFA. The EBA did not care about faces; the FFA did not care about headless torsos. Two specialized regions, two categories, anatomically separable.
The paper was published in Science in September 2001, four years after Kanwisher's FFA paper. The group titled it bluntly: "A cortical area selective for visual processing of the human body."
Where exactly is EBA?
EBA sits in the lateral occipitotemporal cortex (LOTC), at roughly the junction of the middle temporal gyrus and the middle occipital gyrus. In MNI coordinates, the right-hemisphere peak typically falls around x = 47–51, y = −64 to −72, z = −4 to +12 — posterior and lateral, on the outer surface of the occipital-temporal border 2.
In practical terms: if you're looking at a lateral brain image and tracing back from MT+ (the motion area) toward early visual cortex, EBA sits just posterior and slightly inferior to MT+. It is bilateral but right-lateralized, which mirrors the right-hemisphere dominance seen for face processing in the FFA.
Nearby neighbors matter. EBA is adjacent to hMT+ (motion-sensitive), slightly overlaps the lateral occipital area LO (sensitive to object shape in general), and sits close to a region preferentially activated by tools. This neighborhood is not coincidental: bodies move, body parts are manipulable effectors, and the entire cluster of adjacent regions in LOTC may represent different aspects of socially and motorically relevant visual information.
What does EBA respond to?
The short answer is: images of bodies and body parts, excluding faces.
More precisely:
Whole bodies and parts alike. EBA responds to images of full headless bodies and to isolated limbs, hands, and torsos. The response to whole bodies tends to be larger than to isolated parts, but both drive the region above baseline 1.
Static and dynamic bodies. EBA is activated by photographs of still postures as well as by videos of moving bodies. It responds to point-light displays (motion-capture dots arranged as a person walking) more strongly than to scrambled versions of those same displays.
Non-visual body signals. Several fMRI studies found that EBA is activated when participants haptically (via touch) explore body parts — not just when they see them. In one study using congenitally blind participants, EBA showed body-selective responses to haptic exploration, suggesting the region's selectivity for bodies is not purely visual in origin 3. Whether this reflects a genuinely supramodal representation of the body or cross-modal plasticity in early-blind individuals is still actively debated.
Actions involving one's own body. Astafiev et al. (2004) reported that EBA is also activated when participants make goal-directed limb movements — arm or foot movements toward a visual target — even without visual feedback from the moving limb. The region appears to receive motor efference copies or proprioceptive signals alongside visual ones, which puts it at an interface between perceiving other bodies and representing one's own 4.
Causal evidence: what happens when you disrupt EBA?
Correlation between activation and category is suggestive, but the most direct test of any brain region's function comes from disruption studies.
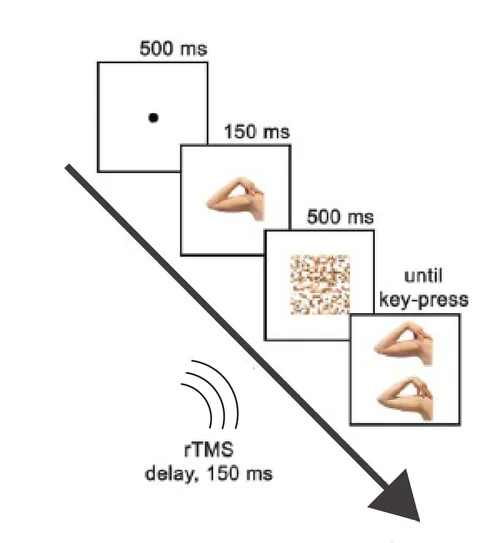
Urgesi et al. (2004) used repetitive transcranial magnetic stimulation (rTMS) to temporarily suppress EBA while participants completed a body-part discrimination task. The task required matching a briefly presented arm or hand to one of two comparison images — identical to the standard discrimination paradigm used for face and object studies.
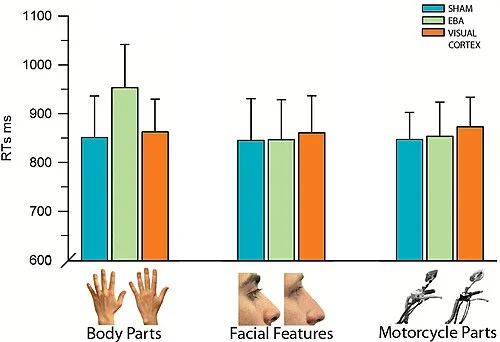
The result was selective: rTMS over EBA increased response times for body-part discrimination by roughly 100 milliseconds compared to sham stimulation. The same TMS pulse train delivered over EBA produced no significant effect when participants discriminated facial features or motorcycle parts. And when TMS was applied over early visual cortex rather than EBA, none of the three category conditions were affected.
A decade later, van Koningsbruggen, Peelen & Downing (2013) extended this logic from isolated body images to real-world scenes 6. In three experiments using fMRI-guided TMS, they asked whether EBA is causally involved in detecting people — not just discriminating body shapes on a blank background, but finding a person among the complex clutter of a street or park scene. TMS over right EBA specifically impaired detection of people in natural scenes, with no effect on detecting cars in the same scenes. Poststimulus TMS (100–200 ms after scene onset) was more disruptive than prestimulus TMS, placing the critical processing window firmly in the early visual response. This was the first direct demonstration that a category-selective region is causally required for detecting its preferred category in natural viewing conditions.
EBA vs. FBA: two body areas, different roles
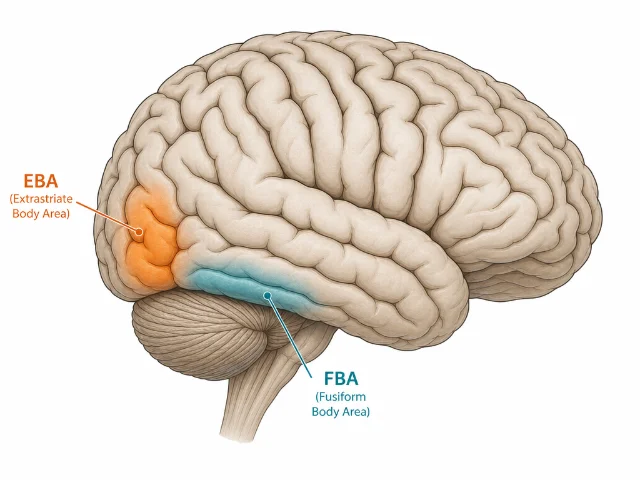
EBA is the more posterior of the two main body-selective areas. A second region — the fusiform body area (FBA) — sits more anteriorly, in the fusiform gyrus, overlapping with or immediately adjacent to the FFA. The two areas have been consistently dissociated:
- EBA responds preferentially to body parts and isolated limbs; it may be primarily engaged in processing the shape and configuration of body segments.
- FBA responds more to whole bodies, is somewhat less sensitive to isolated parts, and shows greater sensitivity to body identity — who the body belongs to.
The functional distinction maps onto an analogy with the face-processing network: EBA may correspond to early form analysis (like the OFA for faces), while FBA may be more involved in identity or whole-body Gestalt processing (like the FFA for faces). This analogy is attractive but imperfect — EBA and FBA were identified by different methods, their developmental trajectories differ, and the depth of their functional dissociation is still being mapped.
What is agreed is that neither area handles faces: damage to the FFA does not impair body recognition, and disrupting EBA leaves face discrimination intact. The double dissociation between body and face processing areas is one of the cleaner boundary-drawing results in human functional neuroanatomy.
Open questions
What exactly does EBA compute? The region is activated by bodies, but the mechanistic description of what it extracts is still debated. Candidates include: the outline or silhouette of body parts; configural relationships between body segments; information about body posture; or something more abstract, like "this visual input belongs to a person." TMS studies that distinguish upright from inverted bodies suggest EBA is sensitive to configuration — it processes inverted bodies differently, though not to the same degree as the FFA handles inverted faces.
How does EBA relate to action understanding? EBA overlaps partially with regions activated by action observation and by the preparation of voluntary movements. Some researchers have proposed that EBA is part of a broader body-representation network that bridges perception and action — a node at which visual body representations and motor body representations interact. Whether this means EBA contains "mirror-like" properties, or simply that it receives convergent projections from motor cortex, is unresolved.
Can EBA be trained or modified by experience? There is evidence that expertise with a body-type modulates EBA responses. Ballet dancers, for example, show stronger EBA activation to dance-posture images than non-dancers. Athletes show body-selective responses modulated by sport-specific body configurations. This suggests the area is not a fixed filter but something that can be calibrated by sustained visual experience with particular body forms.
Is it truly distinct from hMT+? Because EBA sits adjacent to MT+/V5 and both regions respond to biological motion, some authors have questioned whether they are functionally overlapping. Careful parametric studies and retinotopic mapping suggest they are anatomically and functionally separable, but the border region contains voxels with mixed selectivity.
Why EBA matters
The FFA result (1997) established that faces get dedicated cortex. The PPA result (1998) established that scenes do too. EBA (2001) closed a loop: the full human form — not just the face — has its own cortical real estate. This cluster of results together suggests that the brain does not handle visual categories through a single general-purpose object-recognition pipeline. Instead, at least for categories with high behavioral relevance — faces, places, bodies — there is dedicated cortex that handles the specific computational demands of each category.
EBA's existence also raises a question about why. What is so special about bodies that they warrant dedicated processing? One answer centers on the social brain: recognizing the posture, movement, and configuration of other humans is essential for predicting their intentions, understanding their actions, and coordinating behavior with them. A region specifically tuned to body form would give the social brain a faster, more reliable signal about the most behaviorally relevant class of visual objects.
From a broader theoretical perspective, EBA is a test case for the debate between domain-specific and domain-general accounts of high-level visual cortex. Its selective response to bodies, its causal necessity for body discrimination, its anatomical consistency across individuals, and its developmental stability all favor the domain-specific view — that this region is, in some meaningful sense, for processing the appearance of the human body.
Landmark paper: Downing PE, Jiang Y, Shuman M, Kanwisher N. "A cortical area selective for visual processing of the human body." Science. 2001; 293(5539): 2470–2473. PMID: 11577239. DOI: 10.1126/science.1063030.
Course connection: MIT 9.13 covers EBA in the lectures on high-level vision and category-selective regions, as part of the broader series on what visual cortex represents beyond early features. See also Kanwisher's public talks at nancysbraintalks.mit.edu.
Add more perspectives or context around this Post.