Long-Term Depression: How Synapses Learn to Weaken
Long-term depression is the controlled weakening side of synaptic plasticity. This article explains the 1992 Dudek and Bear landmark experiment, the NMDA receptor and AMPA receptor mechanisms behind LTD, why LTD is not just forgetting, and how it complements LTP in learning systems.

If long-term potentiation is the brain's way of turning up a synapse, long-term depression is the equally important act of turning one down. A learning system that only strengthens connections would soon saturate. Every signal would become loud; every association would become sticky. Long-term depression, usually shortened to LTD, gives synapses a way to weaken in a lasting, input-specific way after particular patterns of activity. It is not neuronal damage, fatigue, or forgetting in the everyday sense. It is controlled plasticity in the downward direction.
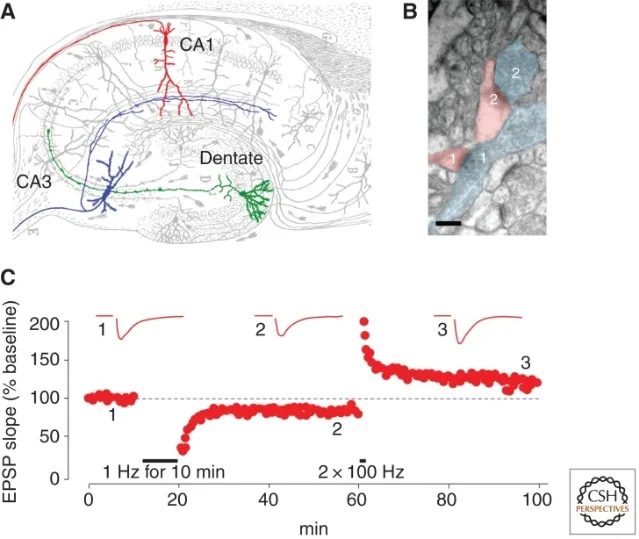
The classic hippocampal version of LTD came into focus in 1992, when Serena Dudek and Mark Bear stimulated the Schaffer collateral input to CA1 neurons in rat hippocampal slices. Their low-frequency protocol, 900 pulses at 1-3 Hz, produced a depression of the CA1 population excitatory postsynaptic potential that lasted for more than an hour after stimulation stopped. The same experiment showed three controls that made the result hard to dismiss: the depression was input-specific, 10 Hz stimulation produced no lasting change, 50 Hz often produced potentiation, and NMDA receptor antagonists blocked the effect. 1
What LTD is
LTD is a long-lasting decrease in synaptic efficacy. At an excitatory synapse, that means the presynaptic neuron can release glutamate as before, but the postsynaptic neuron responds less strongly. The change can last far longer than the stimulation pattern that induced it. Reviews of synaptic plasticity treat LTP and LTD as complementary enduring changes in synaptic strength, both induced by specific patterns of activity and both studied as candidate cellular mechanisms for information storage. 2
That word "specific" matters. LTD is not a whole-cell dimmer switch. In the Dudek and Bear experiment, the depressed pathway weakened while an unstimulated control input did not, which argued against a general loss of postsynaptic excitability. 1 A single CA1 pyramidal neuron receives thousands of excitatory inputs; LTD lets the cell edit some synapses without globally changing its identity.
One useful way to think about LTD is as the answer to a problem posed by Hebbian learning. If "cells that fire together wire together" captures one side of learning, what happens to synapses that repeatedly participate weakly, mistimed, or below threshold? The BCM theory of synaptic modification, developed for visual cortex, predicted a sliding threshold between strengthening and weakening. Bear, Cooper, and Ebner later framed a biologically plausible model in which experience could drive either potentiation or depression depending on activity conditions. 4 Dudek and Bear's hippocampal result gave that prediction a concrete physiological form.
The landmark experiment
The experimental setup was deliberately simple. The researchers cut hippocampal slices, stimulated Schaffer collateral axons from CA3, and recorded the field response in CA1. When they delivered low-frequency stimulation at 1-3 Hz for 900 pulses, the evoked response fell and stayed depressed. When they used other stimulation frequencies, the outcome changed: 10 Hz did not produce a lasting change, and 50 Hz tended to produce LTP rather than LTD. 1
The result was not just "weak stimulation weakens synapses." It was frequency-dependent, pathway-specific, and NMDA receptor-dependent. That combination was the crucial point. A synapse could be driven in one pattern and strengthen; driven in another pattern and weaken. The same receptor family involved in LTP could also help induce LTD.
Dudek and Bear also tested whether the depressed synapses were simply exhausted or damaged. After LTD induction, the same pathway could still support LTP in response to high-frequency stimulation. The synapse had not died; it had changed state. 1
Why the same NMDA receptor can lead to opposite outcomes
The NMDA receptor is often introduced as a coincidence detector. It opens best when glutamate is present and the postsynaptic membrane is depolarized enough to relieve the magnesium block. In LTP, strong coincident activity produces a large postsynaptic calcium signal. In NMDA receptor-dependent LTD, lower-frequency activity can produce a smaller, more prolonged calcium signal. Lüscher and Malenka summarize the broad rule this way: stronger NMDA receptor activation and larger calcium rises favor LTP; more modest, repeated calcium entry favors LTD. 3
That difference in calcium dynamics changes which enzymes dominate. Stronger calcium signals recruit kinase pathways such as CaMKII, which favor AMPA receptor phosphorylation and insertion. Weaker or more prolonged calcium signals can favor phosphatases, which promote AMPA receptor dephosphorylation and removal. 2
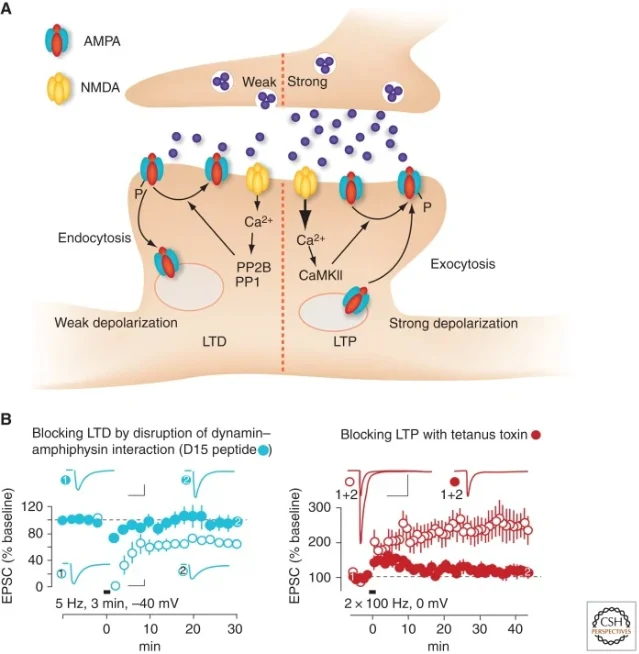
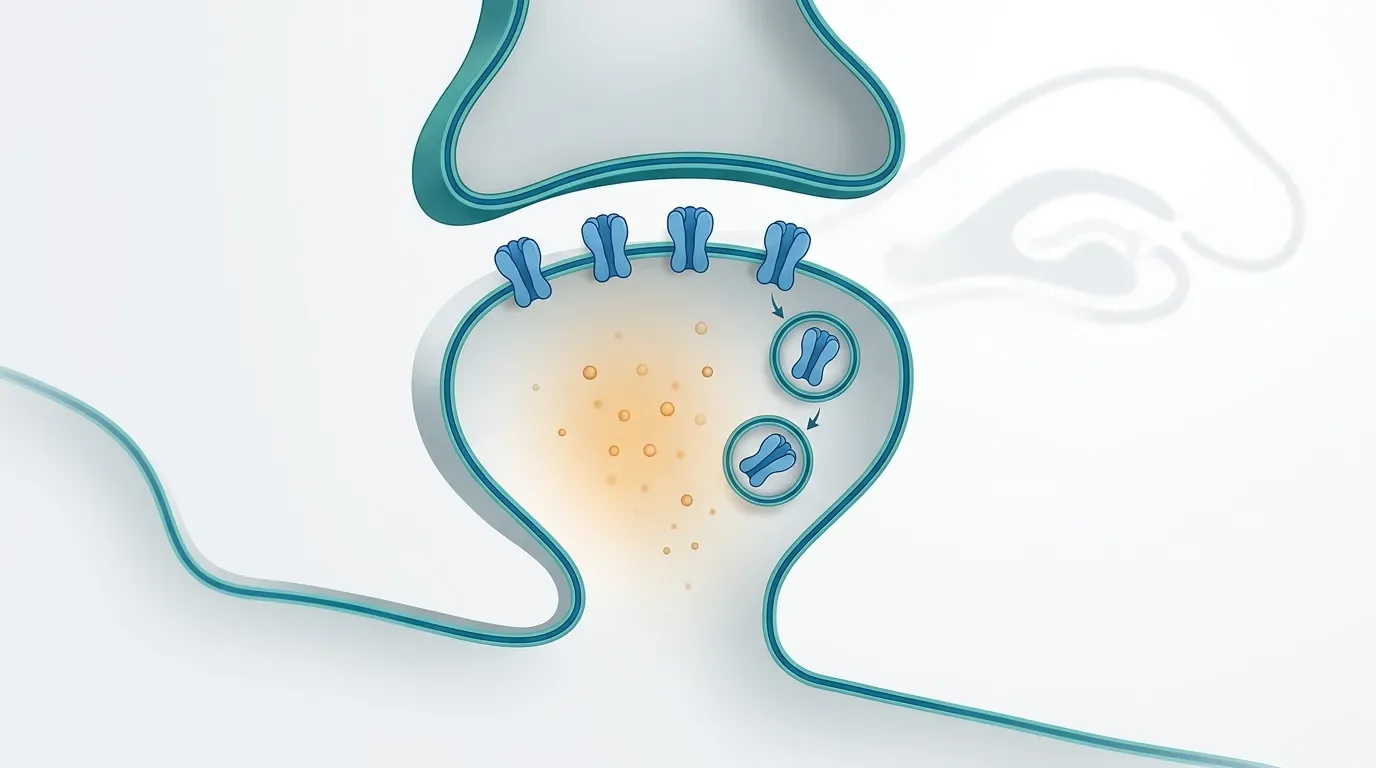
The expression mechanism is often described in terms of AMPA receptors. AMPA receptors carry most fast excitatory current at glutamatergic synapses. During LTP, more AMPA receptors can be inserted into or stabilized at the postsynaptic density. During LTD, receptors can be removed through endocytosis, often from perisynaptic regions followed by redistribution away from the synapse. Blocking endocytosis can inhibit LTD, while blocking exocytosis can abolish LTP. 3
The clean metaphor is volume control, but the real mechanism is inventory control. The synapse changes how many effective receptor channels are available at the membrane and how those receptors are modified.
LTD is not one mechanism
The hippocampal LTD in the 1992 landmark paper was NMDA receptor-dependent. That is only one member of a larger family. Another important form is metabotropic glutamate receptor-dependent LTD, or mGluR-LTD. Gladding, Fitzjohn, and Molnár note that two main forms of hippocampal LTD coexist at CA3-CA1 synapses: one mediated by NMDA receptors and another mediated by metabotropic glutamate receptors. 5
mGluR-LTD is not just the same process with a different receptor label. It can be NMDAR-independent, can recruit group I metabotropic glutamate receptors such as mGlu5, and can involve signaling pathways that regulate AMPA receptor trafficking through MAPK, protein tyrosine phosphatases, and endocytotic machinery. 5 Different brain regions also use LTD differently. Cerebellar LTD, for example, has its own induction logic and is central to theories of motor learning. 2
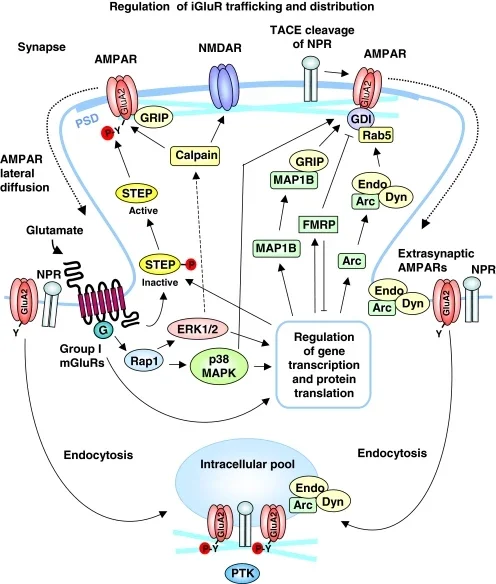
This diversity is one reason LTD can be confusing. The acronym names an outcome, not a single molecular recipe. The outcome is long-lasting synaptic weakening. The route depends on receptor type, cell type, developmental stage, induction protocol, and brain region.
Why LTD matters for memory
LTD matters because a memory system needs selectivity. Strengthening alone can store associations, but it cannot easily remove outdated ones, sharpen contrasts, or keep networks from saturating. LTD offers a way to reduce the influence of synapses whose activity pattern does not match the current computational demand.
This does not mean LTD is "forgetting" in a simple psychological sense. The link from synaptic weakening to behavior is still indirect. Bliss and Cooke stress that whether LTP and LTD are the actual synaptic processes underlying learning and memory has not been definitively resolved, even though plasticity-like changes occur during memory formation and share molecular machinery with memory processes. 2
A more careful statement is this: LTD gives neural circuits a local mechanism for reducing synaptic influence, and that mechanism is plausible enough to belong in any serious account of learning. It may help refine cortical maps, reverse prior potentiation through depotentiation, tune hippocampal representations, and support forms of behavioral adaptation. But no single LTD experiment explains an episodic memory, a learned skill, or the loss of a childhood association by itself.
Open debates
Three debates are worth keeping separate.
First, there is the induction debate. Experimental LTD protocols often use repeated electrical stimulation in slices, such as 1 Hz for minutes. Real brains do not learn by receiving a metronome through an electrode. Spike-timing-dependent plasticity narrows the gap by tying plasticity direction to the relative timing of pre- and postsynaptic spikes, but even that is a simplified window onto behavior. Lüscher and Malenka describe STDP as a more physiological method because such spike relationships may occur during real behavior, not because it fully solves the translation problem. 3
Second, there is the mechanism debate. NMDA receptor-dependent LTD, mGluR-LTD, cerebellar LTD, striatal LTD, and cortical LTD share the name LTD but not all of their molecular machinery. A claim about LTD in one circuit may not carry over to another.
Third, there is the memory debate. LTP and LTD remain powerful cellular models, but memory is distributed across synapses, cells, recurrent circuits, neuromodulators, and systems-level interactions. The right question is not whether LTD "is memory." The better question is when synaptic weakening is the right operation for a circuit trying to learn.
The takeaway
LTD is the brain's controlled way to make a synapse matter less. The Dudek and Bear experiment showed that low-frequency activity could weaken a specific hippocampal input for more than an hour, that the effect depended on NMDA receptors, and that the synapse remained capable of later potentiation. 1 Later work gave the mechanism a molecular vocabulary: calcium thresholds, phosphatases, AMPA receptor trafficking, and parallel receptor systems such as mGluR-LTD.
For a learner, the deepest lesson is bidirectionality. The brain does not merely build stronger connections. It continually edits them.
Landmark paper: Serena M. Dudek and Mark F. Bear, "Homosynaptic long-term depression in area CA1 of hippocampus and effects of N-methyl-D-aspartate receptor blockade," PNAS, 1992. 1
Course connection: MIT 9.13 focuses on how perceptual and cognitive abilities are implemented in the brain and aims to equip students to evaluate empirical articles in cognitive neuroscience. LTD sits below that systems-level curriculum as a cellular mechanism that helps make learning and memory circuits adjustable, including the hippocampal and navigation systems covered in the course. 6 7
Añade más opiniones o contexto en torno a este contenido.