Spike-Timing-Dependent Plasticity: The Brain's Millisecond Learning Rule
Spike-timing-dependent plasticity explains how synapses use millisecond timing to decide whether an input should strengthen or weaken. This article connects the Markram 1997 and Bi & Poo 1998 landmark experiments to NMDA receptor mechanisms, dendritic signals, neuromodulation, and the limits of treating STDP as a universal learning rule.

When two neurons fire close together, the brain does not treat the order as a detail. A spike arriving just before a postsynaptic neuron fires can strengthen a synapse; a spike arriving just after that firing can weaken it. The difference can be a few tens of milliseconds. That is the core idea of spike-timing-dependent plasticity, usually shortened to STDP.
What the rule says
Spike-timing-dependent plasticity is a form of synaptic plasticity in which the relative timing of presynaptic and postsynaptic spikes helps determine whether a synapse undergoes long-term potentiation or long-term depression. In the canonical Hebbian version, a presynaptic spike followed shortly by a postsynaptic spike tends to produce LTP, while the reverse order tends to produce LTD 1.
The intuition is simple. If neuron A fires, and neuron B fires a few milliseconds later, A may have helped drive B. Strengthening A's synapse onto B makes causal sense. If B fires first and A arrives later, A did not help produce that spike. Weakening the connection also makes sense.
This is a sharper version of Hebb's old slogan, often paraphrased as "cells that fire together wire together." STDP adds a harder requirement: together is not enough. Order matters. Timing matters. In many experiments, the decisive window is roughly 20 milliseconds on either side of the postsynaptic spike 2.
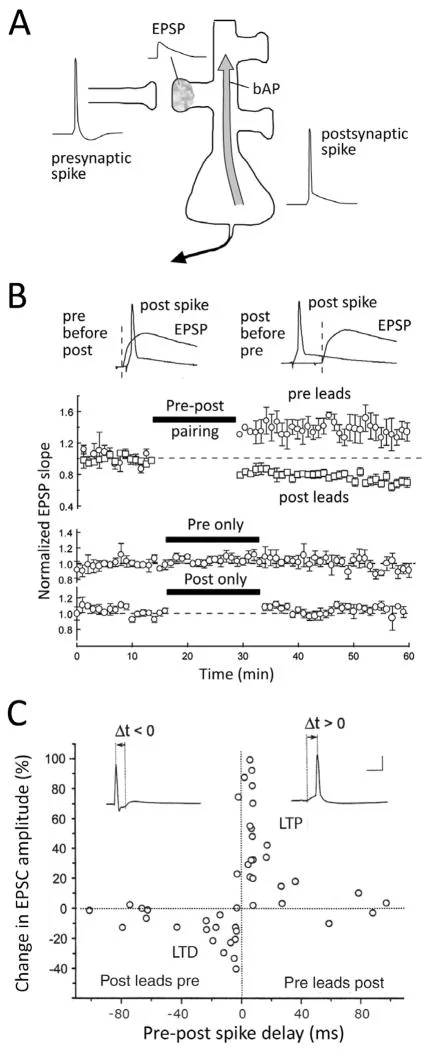
The landmark experiment
The first decisive cortical demonstration came from Henry Markram, Joachim Lübke, Michael Frotscher, and Bert Sakmann in a 1997 Science paper titled "Regulation of Synaptic Efficacy by Coincidence of Postsynaptic APs and EPSPs" 3.
They used dual whole-cell recordings from pairs of layer 5 pyramidal neurons in young rat neocortex. That setup let them control the presynaptic excitatory postsynaptic potential, or EPSP, and the postsynaptic action potential, or AP, with millisecond precision. Bursts of presynaptic spikes alone did not produce a lasting change. Postsynaptic spikes alone did not either. Pairing the two at the right time did 3.
When the postsynaptic action potential occurred about 10 milliseconds after the EPSP, the synapse strengthened. When the postsynaptic action potential occurred about 10 milliseconds before the EPSP, the synapse weakened. When the two events were separated by 100 milliseconds in either direction, the effect largely disappeared 3.
A year later, Guo-qiang Bi and Mu-ming Poo characterized the timing curve in dissociated rat hippocampal neurons. They found LTP when postsynaptic spikes occurred within about 20 milliseconds after presynaptic activation, and LTD when postsynaptic spikes occurred within about 20 milliseconds before presynaptic activation 2. Their experiments also showed that the effect depended on NMDA receptors, that potentiation was strongest at weaker initial synapses, and that the rule did not apply in the same way to glutamatergic synapses onto GABAergic neurons 2.
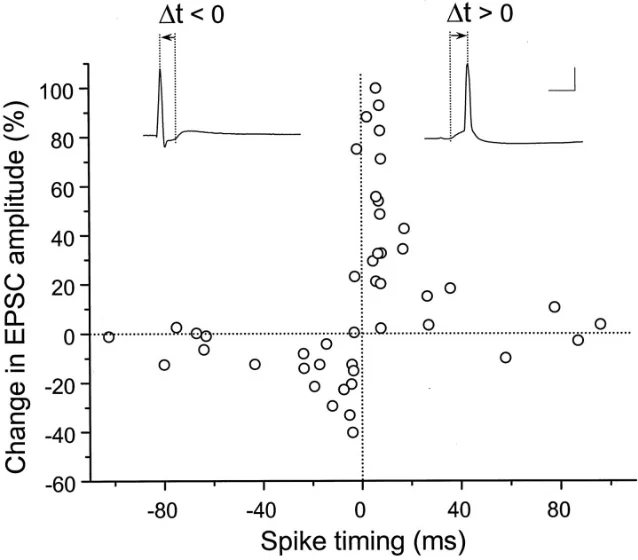
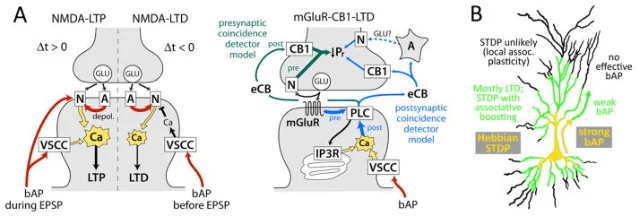
Why milliseconds can change a synapse
At an excitatory synapse, the presynaptic neuron releases glutamate. The postsynaptic membrane responds through AMPA receptors and, if the local voltage is favorable, NMDA receptors. NMDA receptors are useful coincidence detectors because they require both glutamate binding and postsynaptic depolarization to pass calcium efficiently.
In a pre-before-post pairing, the EPSP arrives first and helps depolarize the local dendritic membrane. A back-propagating action potential then invades the dendrite. The two signals overlap, NMDA receptors pass a strong calcium signal, and the synapse is biased toward LTP 1.
In a post-before-pre pairing, the back-propagating action potential has already passed by the time glutamate arrives. The calcium signal is usually weaker or shaped differently, so the synapse is biased toward LTD. In many synapses, LTD can also involve metabotropic glutamate receptors, voltage-sensitive calcium channels, endocannabinoid signaling, or presynaptic changes in transmitter release 1.
The important point is that STDP is not a separate magic rule floating above cell biology. It is one way that ordinary plasticity machinery, NMDA receptors, calcium dynamics, dendritic voltage, AMPA receptor trafficking, and retrograde messengers, becomes sensitive to time.
Why the concept matters
STDP gave neuroscientists a concrete bridge between three levels of explanation.
At the cellular level, it showed how a single synapse can use timing to decide whether an input was plausibly causal. At the circuit level, it helped explain how networks can learn sequences, align spike timing, build receptive fields, and stabilize competition among inputs. In computational neuroscience, STDP became a popular learning rule because it turns temporal order into synaptic change 1.
This is why STDP often appears in models of development and learning. If input A reliably precedes activity in neuron B, the A-to-B synapse strengthens. If another input arrives too late, it weakens. Over many events, the network becomes more sensitive to predictive inputs and less sensitive to lagging ones.
That logic is especially attractive for sequence learning. A chain of neurons that fire in order can, through STDP, strengthen forward connections and weaken reverse or poorly timed connections. The circuit begins to encode the temporal structure of experience, not just which neurons were active.
The rule is real, but it is not universal
The clean cartoon is useful, but the biology is messier. Feldman's review emphasizes that spike timing is one factor inside a multi-factor plasticity rule. Firing rate, dendritic depolarization, synaptic cooperativity, baseline synaptic strength, cell type, developmental stage, and dendritic location can all change the result 1.
Some synapses show Hebbian STDP. Some show anti-Hebbian STDP, where the timing rule is reversed. Some show only LTD. Others express a timing rule only under specific firing frequencies or neuromodulatory states 1. A single postsynaptic spike is not always enough; many protocols require 60 to 100 repeated pairings before a lasting synaptic change appears 1.
That caveat matters because real brains are not quiet slice preparations. In vivo, neurons receive continuous barrages of input while neuromodulatory systems signal attention, reward, novelty, arousal, and behavioral state. Pawlak and colleagues argued that neuromodulators can act as a "third factor" that opens, closes, widens, or shifts the STDP gate 4. Dopamine, noradrenaline, acetylcholine, and other modulators can decide whether a millisecond coincidence is eligible to become a lasting memory trace 4.
The takeaway
STDP is the brain's millisecond-level credit assignment rule. It asks a local question: did this input arrive early enough to help make the postsynaptic neuron fire? If yes, the synapse may strengthen. If no, it may weaken.
That local rule cannot explain memory by itself. It does not replace LTP, LTD, attention, reward, sleep, systems consolidation, or the hippocampal-cortical dialogue. It gives those larger processes a precise synaptic mechanism for one hard problem: turning temporal order into biological change.
Landmark paper: Markram, Lübke, Frotscher, and Sakmann's 1997 Science paper established that the sign of synaptic change could depend on whether the postsynaptic action potential came just before or just after the presynaptic EPSP 3. Bi and Poo's 1998 Journal of Neuroscience paper then mapped the narrow hippocampal timing window in detail 2.
Course connection: MIT 9.13 asks how cognitive abilities are implemented in the brain and trains students to evaluate empirical articles in current neuroscience 5. STDP sits just below that course's systems-level questions: it is one candidate synaptic rule for how experience can alter the circuits that later support perception, learning, and memory.
Añade más opiniones o contexto en torno a este contenido.